[an error occurred while processing this directive]
Post mortem
Elävien lihasten elämä on jatkuvaa
jäykistymistä ja rentoutumista. Kuoleman tultua
lihaksistossa koittaa "viimeinen ponnistus", kun
jäykkyys valtaa koko kehon asteittain. Ilmiö tunnetaan
nimellä rigor mortis, kuolonkankeus.
Rigor mortis on kuolleisiin eläimiin ja ihmisiin
iskevä lihasjännityksen aalto, joka on kaloilla ohi
muutamassa tunnissa, ihmisillä 3-4 vuorokaudessa.
Ihmisillä kuolonkankeus alkaa sydämestä ja
palleasta jo 1/2-2 tunnin kuluttua kuolemasta ja saavuttaa suuret
alaraajojen lihakset 6-8 tunnin kuluttua. Kuolonkankeus vaikuttaa
myös ihmisen ihon karvankohottajalihaksissa, mikä
mikä tekee ruumiin ihosta kananlihamaisen.
Varsinkin ympäristön lämpötila ja liikunto
ennen kuolemaa (ATP:n määrä lihaksissa on
tällöin pienempi) vaikuttavat kuolonkankeuden syntymis-
ja häviämisnopeuteen.
Oikeuslääketieteessä kuolonkankeus
yksinään ei luotettavasti kerro kuolinaikaa, johtuen
juuri olosuhteiden suuresta vaikutuksesta kankeuden
kehittymiseen. Ilmiötä voidaan kuitenkin
käyttää kuolinajan suurpiirteiseen arviointiin ja
kuolinasennon selvittämiseen. Tämä siksi,
että kuolonkankeus kehittyy aikaisemmin ylempänä
olevissa ruumiinosissa.
Kuolonkankeudella on merkitystä etenkin lihan ja kalan
valmistamisessa. Teurastettuja ruhoja riiputetaan, kunnes
kuolonkankeus on ohi. Vasta tämän jälkeen lihaa
voi alkaa valmistaa elintarvikkeiksi. Riiputus nopeuttaa
kankeuden laukeamista venytyksen ansiosta. Kalan kuolonkankeus
kestää selvästi vähemmän aikaa kuin
lihan, mutta esimerkiksi juuri pyydettyjä lohikaloja
kannattaa riiputtaa 1-2 tuntia ennen ruoaksi valmistamista.
Kuolonkankeus heikentää oleellisesti lihan
leikattavuutta ja huonontaa kankeana paistetun kalan mureutta ja
makua.
Kuolonkankeuden lihasjännitystila vastaa normaalissa
lihastyössä syntyvää jännitystä.
Rigor mortiksen syntymekanismin ymmärtämiseksi onkin
ensiarvoisen tärkeää tietää lihaksen
supistumisen ja rentoutumisen biokemialliset reaktiot.
Lihaksen sisin
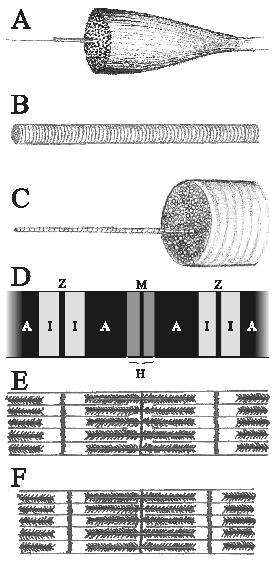 |
Kuva 1: Lihaksen rakenne.
A. Lihas, josta erottuu lihassyykimppu ja edelleen yksi lihassyy.
Kutakin näistä rakenteista ympäröi sidekudostuppi.
B. Lihassyy n. 120-kertaisessa suurennoksessa. Huomaa poikkijuovaisuus.
C. Suurennos lihassyystä, josta erkanee yksi myofibrilli.
D ja E. Myofibrillissä on peräkkäin sarkomeereja. Sarkomeerin alku- ja
loppupäätyä kutsutaan mikroskopiassa Z-linjoiksi. Ohuet aktiinifilamentit
ulottuvat aina puolittain kahden sarkomeerin alueelle. Aktiinisäikeet ovat
isotrooppisia tasopolaroidulle valolle (ks. Seepia 2), joten vaalealla
alueella yksin erottuvia aktiinijuovia sanotaan I-juoviksi. Tummalle
alueelle ulottuvat paksut myosiinifilamentit (A, anisotrooppinen)). H-alue,
jolla on myosiinia mutta ei aktiinirihmoja, näkyy mikroskoopissa harmaana.
-alueen keskellä näkyy M-linja, joka on myosiinifilamenttien keskikohta.
F. Lihassupistuksessa sarkomeeri ja nimen omaan sen H- ja I-alueet lyhenevät.
|
Elektroni-
mikroskooppi avasi tieteenhaaran
Lihassupistukseen liittyy klassinen spiritus animalis-kysymys: mikä antaa eläimelle sen
hengen ja liikkumiskyvyn? Jo
antiikin ajattelijat epäilivät liikkumiskyvyn liittyvän lihaksiin.
Ensimmäiset teoriat lihasten toiminnasta kehitettiin vasta 1930-luvulla. Ne olivat
polymeeritieteeseen perustuvia
ehdotuksia kumimaisesti venyvistä myosiinifilamenteista, joiden liike juontui myosiinin erilaisista
ionisaatiotiloista.
Tämän käsityksen kumosivat kaksi sattumalta samannimistä tutkijaa 1950-luvulla:
Andrew F. Huxley Cambridgesta,
Englannista ja Hugh E. Huxley, Cambridgesta, Massachusettsista. He osoittivat sarkoplasmissa olevan kaksi filamenttia,
jotka liukuivat toistensa lomitse muuttamatta pituuttaan.
Myosiinin ja aktiinin toiminnan selvittäminen molekyylitasolla on ollut ratkaisevasti
ektronimikroskoopin ja sittemmin atomitasolla röntgensädekristallografian ansiota.
|
Lihaskudoksessa on punaisia eli hitaita lihassyitä ja
valkoisia eli nopeita lihassyitä, joiden energiatalous
poikkeaa toisistaan (ks. Maitohappoa ääritilassa, s.
38). Lihassyyn eli lihassolun paksuus vaihtelee välillä
10-100
Lihassyyt yhtyvät pinnastaan ja
päistään sidekudokseen, joka edelleen kiinnittyy
luihin. Lihassupistuksen ja rigor mortiksen mekanismit on
parhaiten selvitetty poikkijuovaisten luustolihasten osalta,
joita siksi käsittelemme tässä lihassoluina.
Lihassyyssä on satoja tai tuhansia fibrillejä,
ohuita säikeitä. Nämä fibrillit
sisältävät puolestaan kukin n. 2500
[1].
Filamentit ovat pitkiä polymeerimolekyylejä,
joita on kahdenlaisia: ohuita aktiinifilamentteja ja paksumpia
myosiinifilamentteja. Nimenomaan aktiini ja myosiini ovat
lihassupistuksen aiheuttajat.
Poikkijuovaisessa lihassyyssä voidaan mikroskoopilla
nähdä raitakuviointi, joka aiheutuu myofilamenttien
sijainnista säännöllisesti vierekkäin. Tummat
ja vaaleat raidat ovat seurausta siitä, että aktiini-
ja myosiiniketjut eivät ole jatkuvia, vaan ovat osittain
lomittain. Siinä, mihin ei ulotu paksuja
myosiinifilamentteja, näkyy siis vain ohuempia
aktiinifilamentteja. Myosiinien kohdat taas
näyttävät tummemmilta. Filamentit ovat oheisen
kuvan mukaisesti lokeroituneet, ja yhtä filamenttilokeroa
kutsutaan sarkomeeriksi.
Myofibrillit kulkevat sarkoplasmassa, joka on lihaksiston
solunestettä. Siinä on runsaasti liuenneena kaliumia,
magnesiumia, fosfaattia ja entsyymejä sekä paljon
mitokondrioita.
Sähkömoottori
Supistuminen ja rentoutuminen ovat monivaiheinen prosessi,
joka alkaa aivojen tai selkäytimen antamalla impulssilla,
käskyllä, joka antaa järjestelmälle nk.
aktiopotentiaalin. Hermosolun ja lihassolun impulsseja
välittävät rakenteet poikkeavat toisistaan,
mikä aiheuttaa sen että lihaksen stimuloimiseen
tarvitaan paljon pidempi jänniteimpulssi kuin hermosolun
stimuloimiseen. Pidemmästä kestosta johtuen impulssien
määrä yhdessä aktiopotentiaalivirrassa voi
olla enintään 100-200. Tämä riittää
kuitenkin aiheuttamaan mahdollisimman suuren supistumisen
sarkomeerissa.
Elvytyksessä sydän käynnistetään
usein antamalla sähköshokki potilaaseen painettavilla
elektrodeilla. Televisiosarjoista tuttu ruumiin nytkähdys
johtuu siitä, että sähköimpulssi vaikuttaa
muuhunkin lihaksistoon kuin sydämeen.
Lihassäikeen keskikohtaan johtaa hermosolun haara,
aksoni, josta hermoimpulssit kulkevat lihassoluun
hermo-lihasliittymän kautta. Siinä lihassolun lomaan
ulottuu aksonin pää, jonka
välittäjäainerakkuloista erittyy
supistuskäskyn tullen kvantittuneesti
("paketteina") asetyylikoliinia. Sen kautta impulssi
kulkee lihassyyn pintakalvoa pitkin kohti syyn molempia
päitä ja edelleen lihassolun sisään
erityisiä kalvoputkia pitkin.
Kalsiumionit aloittavat
|
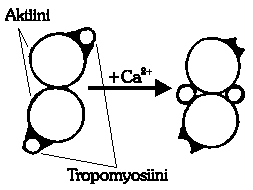
Kuva 2: Poikkileikkaus havainnollistaa
aktiinifilamentin aktiivisten paikkojen vapautumista kalsiumionien
vaikutuksesta.
|
Kalvoputket johtavat myofibrillien pinnalle, missä on
ohutta sarkoplasmaattista kalvostoa. Tällä kalvostolla
on kyky sitoa suuri määrä kalsiumioneja (Ca2+),
mutta myös vapauttaa niitä sarkoplasmin
sisään kalsiumpumppujen kautta.
Kalsiumionikonsentraatio lepotilassa on pienempi fibrillien
sisällä kuin niiden ulkopuolella. Kun kalsiumpumput
tällöin avautuvat aktiopotentiaalin takia, ionit
virtaavat osmoosin ansiosta sarkomeeriin ilman työtä
– ja siis ilman ATP:tä.
Sarkoplasmin sisällä aktiinifilamenttien
ympärillä on kahta säätelyproteiinia:
tropomyosiinia ja troponiinia. Tropomyosiini on aktiinin ja
myosiinin välissä kiinnittyneenä
aktiinipolymeeriin siten, että se estää
myosiinimolekyylin päätä tarttumasta aktiiniin.
Tällöin lihas on rento, sillä kaikki
lihasjännitys aiheutuu myosiinin kiinnittymisestä
aktiiniin. Troponiinia puolestaan on lyhyin välimatkoin
kiinnittyneenä pitkään
tropomyosiiniproteiiniin.
Kalsiumin suuri hetki koittaa: Sarkoplasmaattisesta kalvosta
vapautuu kalsiumioneja sarkoplasmiin. Ca2+ -ionit vaikuttavat
troponiiniin varauksellaan, saaden sen avaruusrakenteessa aikaan
muutoksen. Muutos troponiinissa aiheuttaa puolestaan
rakenteellisen muutoksen tropomyosiinissa, jolloin se muuttaa
asentoaan aktiinijuosteessa (kuva 2). Aktiinin aktiiviset kohdat
paljastuvat, jolloin myosiinimolekyylit voivat kiinnittyä
niihin. Näin huomataan, että kalsiumionien
läsnäolo on "kontraktioliipasin", joka
mahdollistaa vielä huikeamman reaktiokehän...
Energiakysymys
|
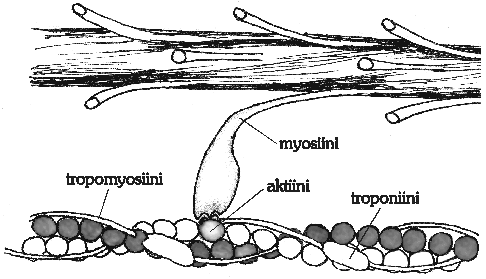
Kuva 3: Myosiinin poikkisilta. Myosiinin kärki
voi kiinnittyä aktiiniketjun yhteen
,
jos pitkä tropomyosiini ja sen seuralainen troponiini ovat väistyneet kuvan 2 mukaisesti.
|
Myosiinimolekyyli jaetaan kahteen osaan. Toinen pää
muodostaa poikkisillan aktiinin kanssa. Myosiinin loppuosa on
pitkä, [a]-kiertynyt ketju, joka liittyy muiden myosiinien
samanlaisten osien kanssa yhteen paksuksi, kierteiseksi kimpuksi.
Näin muodostuu myosiinifilamentti. Kimpun keskikohdasta
myosiinit lähtevät eri suuntiin sarkomeeria, joten
keskellä myosiinifilamenttia on kohta, jossa poikkisiltoja
ei ole. Muissa kohdissa myosiiniketjua raskaat
myosiinimolekyylien päät ulottuvat poikkisiltoina
aktiiniin.
Poikkisilta on kontraktion kannalta mielenkiintoisempi osa.
Sen äärimmäisenä olevaa, paksua
päätyä sanotaan S1:ksi (subfragment-1). S1:n
molekyylirakenne mahdollistaa tarvittavat reaktiot, jotta
aktiini- ja myosiinifilamentit voivat liukua vaiheittain
toistensa ohi.
Kun kalsiumionit ovat "vapauttaneet" aktiinin,
myosiinin pää tarttuu ensiksi aktiinimolekyyliin,
jolloin kompleksi muodostaa 45° kulman. Tällainen
myosiini-aktiiniliitos on hyvin stabiili, ja sitä kutsutaan
-kompleksiksi.
On hyvä ymmärtää, että aktiinin ja
myosiinin sidoksen pysyminen ei kuluta energiaa. Sidoksen tila on
vakaa, ja energiaa tarvitaan lihaksessa vain sidoksen
irroittamiseen.
Energiapotku ja fosfaatti moottoreina
S1-fragmentin molemmissa päissä on aktiiviset
kohdat, joista toiseen voi sitoutua yksi ATP-molekyyli ja toiseen
aktiinimolekyyli. Kuitenkaan ATP ja aktiini eivät voi olla
yhtä aikaa sitoutuneena myosiiniin. Kun nyt myosiini on
rigor-kompleksissa, se on valmis vastaanottamaan toiseen
aktiiviseen kohtaansa ATP-molekyylin, joita on sarkoplasmin
solulimassa.
|
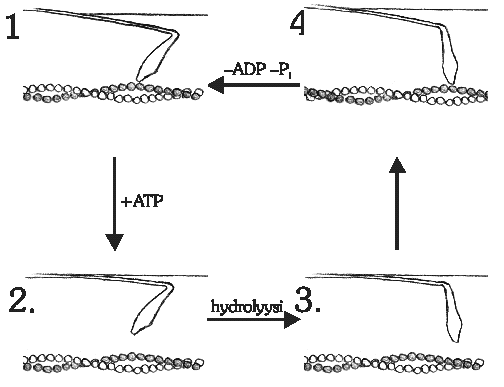
Kuva 5: Poikkisillan askellus. Kuvan mukainen
sykli toistuu aina, kun ATP-molekyyli on myosiinin saatavilla.
1. Rigor-kompleksi. Aktiini (A) ja myosiini (M) ovat kiinni toisissaan.
2. ATP liittyy myosiiniin, jolloin aktiini irtoaa siitä.
3. Tapahtuu ATP:n hydrolyysi, jonka energiapotku kääntää osan
myosiinipoikkisillasta. Tämän jälkeen aktiini voi taas liittyä myosiiniin.
4. ATP:n hajoamistuotteet poistuvat myosiinista, mikä saa myosiinin
S1-fragmentin kääntymään. Kääntyessään S1 liikuttaa aktiiniketjua myosiinin
suhteen.
On palattu asemaan 1.
|
Myosiini on ATPaasi, eli se voi hajottaa ATP:tä.
Tämä onkin luonnollista, sillä ATP
sisältää potentiaalista energiaa myosiinin
käyttöön.
Kun ATP sitoutuu S1:een, se samalla erottaa aktiinin myosiinin
toisesta aktiivisesta päästä. Myosiinin
S1-päässä olevien aktiivisten paikkojen
välissä on näet molekulaarinen
"kuilu", ja ATP:n sitoutuessa myosiiniin, sen
S1-fragmentin osien keskinäiset vuorovaikutukset muuttuvat.
Tämä irrottaa kuilun toisen pään aktiinista.
Kyse on samanlaisesta molekyylistruktuurin muuttumisesta kuin
entsyymien toiminnassa. Myosiini toimiikin reaktiossa osin
entsyymin tavoin, ATPaasina.
ATP:n sitouduttua myosiiniin, rigor-kompleksi on muuttunut nk.
aktiiviseksi kompleksiksi. Tällöin kyseisen
myosiinimolekyylin ja aktiinin välinen jännitys laukeaa
hetkeksi. Kuitenkin, lihastyön aikana
myosiinipäistä on vuorottelevasti kiinni aktiinissa
vain n. 10%, joten koko lihasjännitys ei laukea
samanaikaisesti.
Nyt on myosiinin aika hajottaa ATP hydrolyysin avulla.
Reaktiossa ATP hajoaa ADP:ksi ja epäorgaaniseksi
fosfaattiryhmäksi Pi. Reaktiotyypissä vapautuu kahta
kulutettua ATP:tä kohti 1 vetyioni:
Reaktiossa vapautuva energia on se energia, joka supistaa
lopulta lihasta. S1-fragmentissa aktiinia sitova pää on
selvästi muusta molekyylistä ulkoneva rakenne,
eräänlainen "koura" (engl. "lever
arm"). "Energiapotkun" seurauksena
tämä kohta kääntyy kammen tavoin. Kulman
muutos on 45°. Tällöin myosiinin pää
kääntyy siten, että se muodostaa 90° kulman
aktiinifilamentin kanssa. Myosiinimolekyyli on vielä
irrallaan aktiinista.
Kun myosiini on hajottanut ATP:n, hajoamistuotteet
jäävät kiinni myosiiniin, mutta
myosiinipään S1-rakenne on muuttunut
hydrolyysissä: S1:ssä oleva kuilu on avautunut, ja
aktiini voi jälleen sitoutua myosiiniin.
Aktiinin sitoutumisen myötä fosfaatti (ATP:n
hajoamistuote Pi) poistuu myosiinipäästä (S1).
Fosfaatin poistuminen laukaisee merkittävän
avaruusrakenteellisen muutoksen myosiinipäässä:
S1:n ulkonema ("lever arm") kääntyy
jälleen alkuperäiseen 45 kulmaan, jolloin
päädytään samanlaiseen rigor-kompleksiin kuin
alussa. Ulkoneman kääntyessä aktiini on kuitenkin
tiukasti kiinni myosiinissa, joten ulkonema toimii vipuvartena
kuljettaen aktiinipäätä myosiinin ohi kohti
sarkomeerin keskiosaa.
Nyt rigor-kompleksi on jälleen valmiina reagoimaan uuden
ATP-molekyylin kanssa, joita valmistuu solun
energiantuotantoreaktioissa (ks. Ääritilassa
maitohappoa, viereinen sivu).
Aina lopuksi: rigor mortis
Sidekudoksetkin pilkotaan
Valkuaisaineiden pilkkoutuessa lihaan tulee hyvä, aromikas maku. Myös kestävämmät
sidekudokset ja niiden kollageeni ja elastiini hajoavat, vaikkakin hitaammin kuin myoproteiinit. Kovien sidekudosten hajonneisuus lihassa parantaa kulinaarista kokemusta. Tämän vuoksi naudan lihaa, jossa sidekudosta on enemmän kuin esimerkiksi sian- tai kananlihassa, raakakypsytetään kylmässä (+2°C) ja vakuumissa. Raakakypsytyksessä sidekudosten valkuaisaineet pilkkoutuvat.
Nivelsiteet hajoavat haudassa n. 5 vuodessa, eli niiden hajoaminen kestää 1–2 vuotta
pehmeitä kudoksia kauemmin.
|
Myosiini muodostaa siis aktiinin kanssa kiinteän
rigor-kompleksin, jonka saattaminen aktiiviseksi kompleksiksi
vaatii jatkuvasti uutta energiaa ATP:n muodossa. Lihaksen
supistuminen on jatkuvaa vuorottelua aktiivisen- ja
rigor-kompleksin välillä.
Elävällä eläimellä tai ihmisellä
aktiopotentiaalien sarjaa seuraa aina ennen pitkää se,
että aktiopotentiaalin vapauttama asetyylikoliini tuhoutuu
asetyylikolineraasientsyymin vuoksi, jolloin
aktiopotentiaalivirta loppuu. Tämä saa kalsiumpumput
pumppaamaan kalsiumioneita pois sarkomeerista, jotta lihas voisi
rentoutua ja palautua. Kalsiumionien konsentraation laskiessa
sarkomeerissa alle 6×10-6, aktiinin ympärillä
olevat tropomyosiini ja troponiini estävät jälleen
aktiinia ja myosiinia tarttumasta toisiinsa.
Kuolinhetken tultua tapahtumien kulku ei enää
noudata normaalia, vaan lihassolussa koittaa uudenlainen
tapahtumasarja, joka lopulta johtaa kuolonkankeuteen.
Kuoleman kohdattua, kalsiumpumppujen energiavarastot
ehtyvät ja niiden aktiivinen toiminta lakkaa. Myofibrillien
ulkopuolella sarkoplastisessa kalvostossa
kalsiumionikonsentraatio on paljon suurempi kuin fibrillien
sisällä, missä myosiini- ja aktiinifilamentit
ovat. Kalsiumia alkaa vuotaa sarkoplasmaattisesta kalvostosta
sisälle sarkomeereihin. Kalsiumionit mahdollistavat
myosiinin tarrautumisen aktiiniin, kuten jo tiedämme.
Rigor-kompleksi syntyy. Entä sitten?
Ääritilassa maitohappoa
Punaiset eli ns. hitaat lihassyyt käyttävät energiantuotantoonsa lähinnä rasvahappoja.
Valkeat
eli nopeat lihassyyt taas ovat nimensä mukaisesti suorituskykyisempiä ja ne hapettavat glukoosia.
Glukoosin energian hyödyntäminen alkaa glykolyysillä, jossa glukoosi pilkotaan kahtia.
Samalla
saadaan muutettua kaksi ADP-molekyyliä ATP-molekyyleiksi. Glykolyysiä seuraa lihassolussa usein
varsinkin lihassuorituksen aikana maitohappokäyminen. Se on melko tehoton tapa hyödyntää glukoosin
hajoamistuotteita, mutta maitohappokäymisessä avainsana onkin anaerobisuus. Kun happea on niukasti
saatavilla, glukoosi käy maitohapoksi, joka tosin rasittaa lihassolua. Kaikkein parhaiten glukoosin energiaa voidaan ottaa talteen mitokondrion reaktioissa. Yhdestä glukoosista mitokondrion vetypolttokennoreaktiossa saatava energia on 17-kertainen verrattuna maitohappokäymiseen. Lihassoluissa on paljon mitokondrioita; erityisen paljon niitä on jatkuvasti toimivassa pallealihaksessa ja kaikkein runsaimmin sydänlihaskudoksessa.
Lihassolussa on ATP:hen verrattuna moninkertainen määrä kreatiinifosfaatteja (CP),
joita voidaan
helposti muuntaa ATP:ksi. Esimerkiksi kreatiinitrifosfaatin avulla tuotetaan ATP:tä näin:
|
Lihassolussa on paljon mitokondrioita, mutta aivojen kuollessa
ja verenkierron loppuessa ATP:n tuotanto hiipuu, koska solun
glukoosivarastot ehtyvät eikä toisaalta soluun
enää kulje happea, jota tarvitaan
soluhengityksessä. Jonkin aikaa jatkuu kuitenkin vielä
ATP:n anaerobinen tuotanto maitohappokäymisessä, joka
sekin loppuu happamuuden kasvaessa. Sarkoplasmaan jää
silti vielä ATP:tä.
Kuten muistamme, aktiinin kiinnittyminen myosiiniin
käynnistää myosiinin ATPaasin eli hajotusreaktion.
Reaktion tuloksena rigor-kompleksi laukeaa ja lihas rentoutuu,
mutta vain niin kauan kuin ATP:tä on saatavilla. Kun lopulta
kaikki liikenevä energia on käytetty ja ATP on loppu,
myosiini lukittuu pysyvästi aktiiniin.
Rigor mortis on syntynyt, kuolemasta.
Mätänee, mätänee
Rigor mortiskin häviää lopulta. Elimistön
soluissa käynnistyy hajotustoimintoja,
ensimmäisenä autolyysi. Autolyysissä soluihin
varastoituneet hajottajaentsyymit vapautuvat solulimaan ja
alkavat hajottaa valkuaisaineita.
Autolyysiä seuraa mätäneminen, jonka
aiheuttavat elimistön sisäiset bakteerit ja myös
esim. maaperästä ruumiiseen tunkeutuvat bakteerit.
Mätäneminenkin on entsymaattinen eli entsyymien
katalysoima prosessi, jossa valkuaisaineet hajoavat peptoneiksi
ja edelleen aminohapoiksi, ammoniakiksi, rikkivedyksi, CO2:ksi ja
vedeksi.
Niin hajoavat myös myosiini ja aktiini.
Proteiinifilamenttien särkyessä niiden jännitykset
herpaantuvat ja liha pehmenee jälleen.
Aapo Ahola
Kirjallisuutta
[1] Holmes, K. C.; Muscle Contraction. The
Limits of Reductionism in Biology (ss. 76-92). Novartis
Foundation Symposium 213, 1998.
[2] Guyton, Arthur C.; Textbook of Medical
Physiology (ss. 58-99)
[3] Laiho, Kauno;
Kuolemanjälkeisistä muutoksista.
Oikeuslääketiede (ss. 142-147). WSOY, Porvoo 1970
[4] Contraction, Rigor and Conditioning.
http://www.aps.uoguelph.ca/~swatland/ch5_1.htm
[5] Yann, Brenterch; Olivier, Cazeau;
Romain, Crec’hriou (1997); Transformation du muscle en
viande.
http://www.multimania.com/cazeau/memviande_page4.htm