[an error occurred while processing this directive]
Kummalliset kaksikotiset kukkakasvit
Jos vaikkapa Seepian toimittaja päättäisi
lisääntyä, hän joutuisi etsimään
tarkoitusta varten toisen, eri sukupuolta edustavan yksilön.
Ihmisen sukusolut sisältävät aina 23 kromosomia.
Tämä ei mitenkään riitä elinkelpoisen
yksilön kasvamiseen, eikä itseään voi
siittää.
Kuvitellaanpa vaivattomampi tilanne: toimittaja olisi
kaksisukupuolinen eli ns. hermafrodiitti. Hänelle ei
tuottaisi ongelmia sukupuuton uhatessa laatia itsestään
jälkeläisiä.
Kukkivista kasveista suurin osa on hermafrodiitteja. Kaikki
hermafrodiitit eivät kuitenkaan pölytä helposti
itseään. Itse asiassa ne voidaan karkeasti jakaa
kahteen suureen ryhmään, ristipölyttäjiin ja
itsepölyttäjiin. Vain harvat kukkakasvit tuottavat
yhtä paljon sisä- ja ristisiittoisia
jälkeläisiä.
Itsepölytteisyys
Itsepölytteisyys on harvoin poikkeuksetonta. Esimerkiksi
papu on pääasiassa itsepölytteinen, mutta tuottaa
osan jälkeläisistään
ristipölytyksellä. Äärimmäisen
itsepölytteisiä ovat monet ns. kleistogamiset
orvokkilajit. Kleistogamia merkitsee kukan avautumista vasta
pölytyksen jälkeen.
Itsepölytyksessä jälkeläisen geneettinen
materiaali köyhtyy, koska osa emokasvin geeneistä
jää periytymättä. Emokasvilla saattaa olla
myös jokin peittyvä tai muuten harmillinen
geeni yhtenä kappaleena, jolloin sen vaikutus ei näy.
Tämä geeni saattaa periytyä sekä naaras-
että mukana kaksinkertaisena
jälkeläiselle, joka tällöin kuolee tai
ainakin kärsii. Ilmiötä kuvaa englanninkielinen
termi inbreeding depression, suomeksi sisäsiittoisuuden
aiheuttama heikentyminen. On huomattava, että
itsepölytteisten lajien genomi on jatkuvan
itsepölytyksen takia ylipäänsä köyhempi
kuin DNA:n määrästä voisi
päätellä.
Toisaalta itsepölytys tarjoaa kasville keinon
lisääntyä silloinkin, kun muita saman lajin
yksilöitä ei ole lähettyvillä. Tämä
on suuri etu mm. kolonisaation eli uusille alueille
levittäytymisen kannalta. Mikäli
ristipölytystä kuitenkin tapahtuu, ei geneettisen
muuntelun ja siten runsaan sopeutuvaisuuden mahdollisuutta ole
ratkaisevasti rajoitettu.
Apomiksia ja kasvullinen lisääntyminen
Esimerkiksi monet heinät, voikukat ja karhunvatukat ovat
apomiktisia lajeja. Toisin sanoen ne tuottavat siemeniä
ilman hedelmöitystä. Tällöin kromosomisto ei
puolitu sukusolun syntyessä, ja jälkeläinen on
emokasvin klooni. apomiksia esiintyy usein yhtäaikaa
suvullisen lisääntymisen kanssa. apomiksia ei
tietenkään salli geneettistä muuntelua, mutta ei
myöskään johda sisäsiittoisuuden haittoihin.
Kasvulliseen lisääntymiseen (esimerkiksi rönsyjen
ja maavarsien avulla) verrattuna apomiksilla on se etu, että
siemenet voivat levitä kauas uusille kasvupaikoille.
Itsepölytteisyyden tavoin on apomiksia tehokas
lisääntymistapa stabiilissa
ympäristössä, johon kasvi on hyvin sopeutunut.
Kaksikotisuus ilmiönä
Toiset kasvit ovat vähemmän kärkkäitä
tuottamaan siemeniä ilman geneettistä muuntelua.
Itsepölytystä voi ehkäistä eriaikaisuus,
heterostylia tai muu kukan rakenteellinen ominaisuus,
yksikotisuus, itseinkompatibiliteetti tai kaksikotisuus (katso
sivu 6). Eläimillä kaksikotisuus on
äärimmäisen yleistä, mutta kasveista
kaksikotisina pidetään erilaisten arvioiden mukaan vain
noin 2-12 prosenttia (n. 6% eli n. 14 620 lajia, Renner &
Ricklefs [9]).
 |
|
Kuva 1:Hillan (Rubus chamaemorus)
emikukkia (yllä) on vain noin puolet hedekukien (alla)
määrästä. Verho- ja terälehtiä
kukissa on yleensä viisi, mutta joskus myös neljä
tai kuusi. Hedelehtiä kukissa on yleensä 25 – 120
ja emilehtiä 3 – 35. Kukkien tärkeimpiä
pölyttäjiä ovat kukkakärpäset.
|
 |
Tarkan osamäärän ilmoittamista vaikeuttaa
paitsi se tosiasia, että kaikkia maailman kasveja ei
suinkaan täysin tunneta, myös kaksikotisuuden
määrittelyyn liittyvät ongelmat. Suuri joukko
kasveja lisääntyy jollakin kaksikotisuuden kaltaisella
mekanismilla (esim. gyno- ja androdieekkisyys, katso sivu 6).
Hyvin yleistä on myös seksuaalinen epävakaus ja
sukupuolen vaihto, jolloin jomman kumman tai joskus jopa
molempien sukupuolten yksilöt saattavat vaihtaa sukupuolta
kokonaan tai osittain tai tuottaa hermafrodiittisia kukkia.
Seksuaalisen epävakauden ilmenemiseen vaikuttavat
esimerkiksi energiankulutus ja ravinnetilanne.
Jotkut lajit ovat toiminnallisesti kaksikotisia, vaikka
kukissa olisikin ylimääräisiä
sukupuolielimiä. Esimerkiksi tietyillä viikunalajeilla
”hedeyksilöiden” emit eivät tuota
siemeniä, vaan tarjoavat symbioottisille
pölyttäjähyönteisille paikan munia.
Emiyksilöissä taas ei ole heteitä lainkaan.
Miksi valita kaksikotisuus?
Voidaan kysyä, miksi kaksikotisuus on näinkin
harvinainen piirre kasvikunnassa. Jos verrataan sitä muihin
sisäsiittoisuudenestomenetelmiin, havaitaan niiden olevan
monin tavoin edullisempia. Eriaikaiset, heterostyliset ja
itseinkompatibiilit kukat kykenevät periaatteessa jokainen
tuottamaan hedelmän, kun taas kaksikotisen lajin hedekukat
eivät tähän pysty. Lisäksi esimerkiksi
itseinkompatibiliteetin ei tarvitse olla täydellistä,
jolloin sisäsiittomahdollisuus takaisi kasville
lisääntymismahdollisuudet kaukana lajitovereistakin.
Edelleen hermafrodiitissa kukassa yhdet pölyttäjiä
houkuttelevat elimet palvelevat sekä emien että
heteiden tavoitteita toisin kuin hede- tai emikukassa.
Esitämme kysymyksen toisin päin: miksi ihmeessä
olla kaksikotinen? Entä mikä yhdistää
kaksikotisia lajeja?
 |
|
Kuva 2:Asterikasveihin kuuluvan
kaksikotisen kissankäpälän (Antennaria dioica)
emikukat ovat valkoisia ja hedekukat vaaleanpunaisia. Kasvi on
yleinen Suomessa valoisilla ja kuivilla paikoilla.
|
Kaksikotiset kasvit ovat pääasiassa monivuotisia.
Tämä tuntuu ymmärrettävältä:
monivuotisella kasvilla ei ole kiirettä kukkia ja tuottaa
jälkeläisiä heti, vaan se voi odotella
onnistunutta pölytystä vuosikausia. Monivuotisen kasvin
geneettinen muuntelu on sitä paitsi yksivuotista
vähäisempää pitkän eliniän takia.
Tämän voidaan ajatella asettavan korkeita vaatimuksia
muuntelulle silloin kun sitä pääsee
tapahtumaan.
Myös kasvin koko on suoraan verrannollinen
kaksikotisuuden esiintymismäärään. Suuren
yksilökoon ajatellaan johtavan helposti itsepölytykseen
kasvin omien kukkien kanssa. Tällöin
sisäsiittoisuutta pyrittäisiin jollain keinolla
välttämään.
Kaksikotisuus on yleisintä tropiikissa ja harvinaistuu
napoja kohti. Tämän oletetaan johtuvan monivuotisten
kasvien (varsinkin puiden; trooppisista puista huomattava osa on
kaksikotisia) yleisyydestä lämpimillä seuduilla.
Onhan kylmä talvi monivuotiselle kasville melkoinen rasite.
Toinen selitys ilmiölle liittyy kaksikotisten kasvien
siementen leviämistapaan: kaksikotiselle kasville
tyypillinen piirre on mehevien hedelmien tai marjojen
tuottaminen. Tällainen hedelmä houkuttelee lintuja
ruokailemaan ja samalla levittämään siemeniä.
Tropiikissa siementen leviäminen lintujen
välityksellä on huomattavasti tehokkaampaa kuin
kylmemmillä ilmastovyöhykkeillä.
Pölytys
Kun tutkitaan, millaisia ominaisuuksia on samoihin
kaksikotisten kasvien kanssa kuuluvilla lajeilla,
löydetään voimakkaita korrelaatioita mm.
kaksikotisuuden, yksikotisuuden ja ns. abioottisen
pölytyksen välillä. Tätä
ilmiötä voimme yrittää selittää
pölytystavan ominaisuuksilla: tuuli ja vesi kuljettavat
siitepölyä sattumanvaraisesti, eläimiä varten
kasvi taas pystyy laatimaan sen muotoiset
lisääntymiselimet, että siitepöly
päätyy suurella todennäköisyydellä
pölyttäjän kyytiin ja siitä edelleen
seuraavaan kukkaan. Abioottisen pölytystavan siis oletetaan
kuljettavan huomattavan osan kukan tuottamasta
siitepölystä sen omiin sukuelimiin. Tämä
aiheuttaisi itsepölytyksen tai itseinkompatiibelissä
kukassa ehkä 'tukkisi' emit vieraalta
siitepölyltä.
Yksi- ja kaksikotisuus ovat keskittyneet pikemmin
primitiivisiin kuin pitkälle erikoistuneisiin taksoneihin
ainakin kaksisirkkaisissa kasveissa. Tämä sopii yhteen
edellä mainitun kanssa: pitkälle kehittyneiden
kukkakasvien kukkien rakenne on usein tarkasti
määritelty ja pölyttäjien toimintaa
suosivaksi muotoiltu.
|
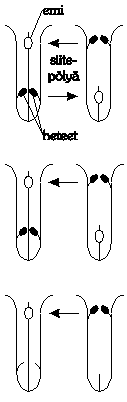
Aivan erikoislaatuinen tapauskohtainen selitys
kaksikotisuudelle on Beachin ja Bawan hypoteesi heterostylian
evoluutiosta kaksikotisuudeksi.
Alkutilanteessa heterostylisen kukan
pölyttäjä on erikoistunut
”pitkäkärsäinen” hyönteinen, joka
kuljettaa siitepölyä kumpaankin suuntaan.
Oletetaan, että jossain kasvin
esiintymisalueella hyönteisfauna muuttuu siten, että
”pitkäkärsäiset” hyönteiset
kuolevat pois. Uudet ”lyhytkärsäiset”
pölyttäjät eivät ulotu kukan alaosiin asti,
jolloin siitepöly kulkee vain yhteen suuntaan.
Tällöin heterostylisen kukan ”matalat” emit
ja heteet surkastuvat vähitellen pois tarpeettomina.
|
 |
Kenties myös geneettinen tie yksisukupuolisia kukkia
tuottavasta yksikotisesta lajista kaksikotiseen on vaivattomampi
kuin hermafrodiittikukkia tuottavasta lajista. Kaksikotisuus
voisi heijastaa esim. siitepöly- ja siementuotannon
kannattavuutta erilaisilla kasvupaikoilla, niin että tietyt
kasvupaikat olisivat suosineet jommankumman sukupuolen kukkien
vähenemistä yksilössä.
Hyönteispölytteisten kaksikotisten kasvien kukat
ovat tavallisesti tuulipölytteisten kasvien kukkien
tapaisia: pieniä, yksinkertaisia ja vaatimattoman
värisiä. Vaatimaton voimavarojen käyttö kukan
muodostamiseen sopisi hyvin yhteen sen tosiasian kanssa,
että kukkalehdet palvelevat kaksikotisuudessa
”epätaloudellisesti” vain yhtä sukupuolista
tehtävää yhdessä kukassa.
Näyttävät ja monimutkaiset kukat ovat
tunnusomaisempia hermafrodiittilajeille. Näissä
käyvät pölyttäjät ovat usein
erikoistuneita, ja kukka saattaa rakenteellaan
”valita” haluamansalaisen pölyttäjän.
Erityisen selvä tehtävä pölyttäjän
anatomialla on monissa heterostylisissä kasveissa.
Kaksikotisuuden on esitetty liittyvän
'erikoistumattomiin' hyönteisiin, joilla ei ole
mihinkään kukkarakenteeseen sopeutunutta
ruumiinrakennetta ja jotka (ehkäpä juuri siksi)
viettäisivät yhdessä kukassa möyrien
kauankin. Kaksikotisuudella laji saa aikaan riittävän
siitepölyvirran yksilöiden välille. Kun
hedekukassa emit eivät vangitse omaa siitepölyä
eikä emikukassa oma siitepöly tuki emejä, voi
järjestelmä olla edullinen itseinkompatibiliteettiin
nähden.
Toinen hypoteesi liittyy tiettyjen mehiläisten
hyvään muistiin ja mahdolliseen
pesänsisäiseen kommunikaatioon runsaiden ruoka-apajien
sijainnista. Jos hyönteiset ryhtyvät ahkerasti
käymään hermafrodiittikasvin harvinaisen
runsaskukkaisessa yksilössä ja vasta tämän
ravinnonlähteen ehtyessä siirtyvät
vaatimattomampiin yksilöihin, liikkuu siitepöly
lähinnä ensimmäisistä kukista
jälkimmäisiin. Tätä voisi seurata
houkuttelevampien yksilöiden keskittyminen hedetoimintaan ja
vaatimattomampien siementuotantoon.
Yleensä kaksikotisten kasvien hede- ja emikukkien suhde
on epätasainen: emikukkia on tavallisesti
vähemmän. Hillalla, joka on Suomessa yleinen
kaksikotinen monivuotinen kasvi, emikukkia on vain noin kolmasosa
kaikista kukista. Hillan pölyttäjät ovat
nimenomaan edellämainitun kaltaisia erikoistumattomia
pölyttäjiä. Hillan emikukissa ei ole juuri
lainkaan mettä, mikä antaa aihetta olettaa, että
pölyttäjät käyvät niissä vain
'vahingossa', ulkonäön samankaltaisuuden vuoksi. Monien
muidenkin hyönteispölytteisten kaksikotisten kasvien
oletetaan 'huijaavan' pölyttäjiä samalla
tavoin.

|
|
Kuva 4:Kaksikotinen
kanariantaatelipalmu (Phoenix canariensis) on läheistä
sukua varsinaiselle taatelille (Phoenix dactylifera). Sen
hedelmät ovat syötäviä, mutta eivät
yhtä hyviä kuin oikeat taatelit.
|
Resurssiallokaatio
Myös voimavarain käyttöön liittyvät
syyt yhdistetään kaksikotisuuden evoluutioon.
Hermafrodiittikukkia tuottava yksilö joutuu jakamaan
käytettävissä olevat varat kahden sukupuolisen
tehtävän välillä. Kaksikotinen yksilö
taas voi keskittyä omaan
lisääntymistehtäväänsä.
Tarkastelemme tätä ns. resurssiallokaatiota
lajinsisäisen kilpailun kautta. Siitepölyn tuottajana
kasvin optimaalinen lisääntyminen merkitsee
siitepölyn levityksen maksimointia. Siementämisen
onnistumisen taas ratkaisee siemenen ja hedelmän tuottoon
käytettävissä olevien resurssien
määrä. Tällöin siitepölyn tuottajan
etu on maksimoida pölytysten määrä.
Mikäli siitepölyn leviäminen paranee
riittävästi yksilön investoidessa
pölyttäjien houkutteluun, sen kannattaa
jättää emien tuotanto sikseen.
Jos vastaavasti siementen leviäminen paranee paljon
toisten yksilöiden keskittäensä voimansa
levittäjiä houkuttelevan hedelmän luomiseen, on
näiden syytä unohtaa heteet. Tämä malli sopii
mainiosti yhteen edellä esitetyn yksisuuntaisen
siitepölyn kulun kanssa.
Edellisen pohdinnan valossa huomiota herättää
esimerkiksi Steve Sutherlandin havainto, jonka mukaan
itseinkompatibiileista hermafrodiittikukista tuottaa
hedelmää vain n. 22%. Tämä ei poikkea
paljoakaan kaksikotisen kasvin hedelmien ja kukkien
välisestä määräsuhteesta. Eikö
tämä tue kasvien sukupuolista erilaistumista siinä
tapauksessa, että yksilö voisi jotenkin vaikuttaa
siihen, kumpi seksuaalinen tehtävä sille olisi
optimaalinen?
Muita selityksiä
Geneettisesti kaksikotisuuden kehittyminen on paljon
vaivattomampaa kuin
itseinkompatibiliteettijärjestelmän. Jo pienet
hormonaaliset muutokset voivat saada aikaan sukupuolen
vaihtumisen.
Myös tuholaisten torjuminen voi joissakin tapauksissa
johtaa kaksikotisuuteen. Erikoinen esimerkki tästä on
edellä mainittu viikunan funktionaalinen kaksikotisuus,
jossa ”vale-emikukat” on sijoitettu optimaalisesti
pölyttäjien tyyssijoiksi; nähtävästi
yksikotisen viikunan toisenlainen kukkimistapa on tietyissä
tuholaisfaunoissa altis hyödyttömille emeihin muniville
hyönteisille. Joidenkin kaksikotisten lajien tuholaiset ovat
erikoistuneet hedekukkasilmuihin. Vaikka heteitä
tällöin tuhoutuisikin huomattavasti, saattaa haitta
olla vähäisempi kuin vastaavasti hermafrodiittisilmun
tuhoutuessa, koska vain toinen lisääntymistoiminto
häiriintyy ja siemeniä tuottava kukka on kuitenkin
jäänyt koskematta.
Seksuaalinen kaksimuotoisuus ja segregaatio
Kaksikotisilla kasveilla yleistä on seksuaalinen
kaksimuotoisuus ja sijoittuminen erilaisille kasvupaikoille, ns.
segregaatio. Näiden voidaan ajatella heijastelevan eroja
kasvien resurssienkäytössä. Rajoituksensa
näille ominaisuuksille asettaa se, että kummankin
sukupuolen on kuitenkin käytettävä samaa
pölyttäjää. Voi olla, että
kaksimuotoisuus ja
segregaatio toimivat välineinä pölyttäjien
kulkusuunnan määrittämisessä (hedekukkien
preferoimiseksi, vrt. hillan emikukat aikaisemmin
tekstissä).
Kuinka kaksikotinen kasvi kolonisoi?
Trooppisten saarten kasvistoa tutkittaessa on havaittu,
että kaksikotiset kasvit pystyvät
todennäköisesti valtaamaan uusia, kaukaisia
kasvupaikkoja eli kolonisoimaan suurin piirtein yhtä
tehokkaasti kuin hermafrodiittiset ja apomiktisetkin lajit.
Tämä tuntuu yllättävältä ottaen
huomioon kaksikotisen kasvin hankalan siementuoton: ilman
erisukupuolista lajitoveria ja onnistunutta pölytystä
yksilöiden välillä siemeniä ei synny. Voidaan
kuitenkin esittää seuraavia syitä kaksikotisten
kasvien pärjäämiselle:
 |
|
Kansikuva: Kataja on tyypillinen
esimerkki kaksikotisesta kasvista. Emiyksilöihin (kuvassa)
syntyy pieniä marjamaisia käpyjä, joiden
kypsyminen kestää kaksi vuotta.
|
1) Kaksikotisuus on harvoin ehdotonta. Yksikotisia ja
hermafrodiitteja yksilöitä esiintyy hyvin monissa
kaksikotisissa lajeissa. Nämä poikkeusyksilöt
eivät yleensä ole itseinkompatibiilejä. Kenties
näillä harvinaisilla erikoistapauksilla on olennainen
ekologinen tehtävä kolonisaatiossa. Toinen selitys
nimenomaan sellaiselle seksuaaliselle epävakaudelle,
että emikukat joskus tuottavat heteitä, olisi
pölyttäjien houkuttelu emikukkiin. On lajeja, jotka
tuottavat tähän tarkoitukseen hermafrodiitin
näköisissä emikukissa steriiliä
siitepölyä.
2) Kaksikotisten kasvien yleinen leviämistapa marjojen
välityksellä voi auttaa asiaa: siementen
levittäjä (yleensä lintu) syö kerralla kaikki
marjassa olevat siemenet ja kuljettaa ne samalle seudulle.
Tällöin kolonian muodostavat todennäköisesti
molempien sukupuolten edustajat. On huomattava, että
lähes kaikki kaksikotiset sammalet levittävät
itiönsä neljän kappaleen könteissä,
joissa on kaksi ”koiras-” ja kaksi
”naarasitiötä”.
3) Monet kaksikotiset kasvit ovat sitkeitä ja
monivuotisia. Ne myös kasvattavat voimakkaasti esim.
maavarttaan ja lisääntyvät suvuttomasti laajalle
levittäytyneen yksilön katkeillessa. Kasvit voivat
siten odotella pölytystä vaikkapa kymmeniä vuosia.
Esimerkiksi hilla kasvattaa pitkiä maavarsia, jotka
katketessaan palasiksi muodostavat usean geneettisesti identtisen
yksilön kloonin. Hillan suvullinen lisääntyminen
on nykyisissä ekologisissa oloissa
käytännöllisesti katsoen
merkityksetöntä, ja hillakloonien iät ovatkin
tuhansia vuosia.
Jaakko Kortesharju
Kirjallisuutta
[1] Baker, H. G. & Cox, P. A. 1984.
Further thoughts on dioecism and islands. Annals of the Missouri
Botanical Garden 71: 244-253.
[2] Barrett, S. C. H. 1984. Variation in
floral sexuality of diclinous Aralia (Araliaceae). Annals of the
Missouri Botanical Garden 71: 278-288.
[3] Bawa, K. S. 1980. Evolution of dioecy in
flowering plants. Annual Review of Ecology and Systematics 11:
15-39.
[4] Bawa, K. S. 1984. The evolution of
dioecy - concluding remarks. Annals of the Missouri Botanical
Garden 71: 294-296.
[5] Beach, J. H. 1981. Pollinator foraging
and the evolution of dioecy. The American Naturalist 118:
572-577.
[6] Charnov, E. L., Smith, J. M. & Bull,
J. J. 1976. Why be an hermaphrodite? Nature 263: 125-126.
[7] Freeman, D. C., Lovett Doust, J.,
El-Keblawy, A., Miglia, K. J. & McArthur, E. D. 1997. Sexual
specialization and inbreeding avoidance in the evolution of
dioecy. The Botanical Review 63: 65-92.
[8] Haber, W. A. & Bawa, K. S. 1984.
Evolution of dioecy in Sarauia (Dilleniaceae). Annals of the
Missouri Botanical Garden 71: 289-293
[9] Renner, S. & Ricklefs, R. 1995.
Dioecy and its correlates in the flowering plants. American
Journal of Botany 82(5): 596-606
[10] Sutherland, S. 1986. Floral sex ratios,
fruit-set, and resource allocation in plants. Ecology 67:
991-1001.
[11] What is apomixis? (1998).
http://billie.harvard.edu/apomixis/apomixis.html
[12] Genetics 364 Lectures – Lecture
2: Breeding Systems in Plants.
http://www.biology.ualberta.ca/courses.hp/gen364/ag_2.html
[13] Sex determination and
self-incompatibility.
http://www.css.orst.edu/CLASSES/CSS530/Unit15.htm
[14] Malcomber, S. (1996); Gaertnera
(Rubiaceae) – Using phylogenies to study the evolution of
breeding systems.
http://www.mobot.org/MOBOT/research/gradstudents/malcomber/gaertweb.html
[15] Evolution of Self-Fertilization.
http://biology.queensu.ca/~biol206/selfing.htm
[16] Husband, B. (1999); The Functional
Significance of Dichogamy in Chamerion angustifolium.
http://www.uoguelph.ca/botany/evollab/dichogamy.htm
[17] Won, H. & Renner, S. (2000); Why so
many dioecious species in Siparunaceae (Laurales)?
http://www.botany2000.org/section13/abstracts/146.shtml
[18] Garbutt, K.; Inbreeding depression in
plants and its consequences.
http://www.as.wvu.edu/~kgarbutt/Papers2/Inbreed.html