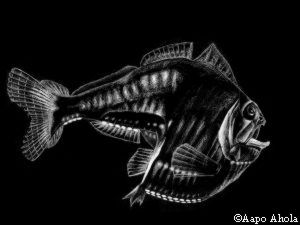
Kuva 1: Syvänmerenhelmiäiskala (Argyropelacus aculeatus) sivulta (vasemmalla) ja alhaaltapäin (oikealla). Valoelinten tuottama valo auttaa kalaa sulautumaan veden pinnan läpi kuultavaan auringon valoon.
Madeiran saarella kalastajat houkuttelevat kaloja valoa tuottavien kalmarien paloilla, joita he kiinnittävät pitkien siimojensa päähän. Kalmarien tuottama valo on kylmää. Se on peräisin kemiallisesta reaktiosta, jonka sivutuotteena ei synny juuri lainkaan lämpöä. Valon kemiallista syntymistä elävissä organismeissa kutsutaan bioluminesenssiksi.
Bioluminesenssia tavataan lähes kaikissa eliöryhmissä, kuten bakteereissa, levissä, sienissä, alkueläimissä, hyönteisissä, nilviäisissä ja kaloissa. Valoa tuottavat eliöt kuitenkin puuttuvat esimerkiksi putkilokasveista ja maalla elävistä selkärankaisista. Suurin osa valoa tuottavista eliöistä elää meressä.
LuminesenssiLuminesenssi tarkoittaa tapahtumaa, jossa atomin tai molekyylin viritystila purkautuu, ja atomi palaa alemmalle energiatasolle vapauttaen ylimääräisen energian valona. Jos luminesenssi tapahtuu heti virittymisen jälkeen, sitä kutsutaan fluoresenssiksi. Pitkän ajan päästä tapahtuvaa luminesenssia kutsutaan fosforenssiksi. Viritystilan aiheuttaneen energialähteen mukaan luminesenssi luokitellaan seuraavasti: [14] FotoluminesenssiFotoluminesenssissa atomiin osuva fotoni virittää sen. Viritystilan purkautuessa voi syntyä joko samanlainen fotoni, kaksi tai useampia fotoneita, joiden yhteenlaskettu energia on yhtä suuri kuin alkuperäisen, tai pienempienerginen fotoni, jolloin osa energiasta muuttuu lämmöksi. Fotoluminesenssia esiintyy muun muassa fosforipitoisissa kivissä ja pimeässä loistavissa tarroissa (fosforenssi) sekä hampaissa ja monilla valkaisevilla pesuaineilla pestyissä vaatteissa, kun niitä tarkastellaan ultraviolettivalossa. KemiluminesenssiKemiallisessa reaktiossa syntyvän reaktiotuotteen konfiguraatio ei ole optimaalinen, jolloin se hetken kuluttua muuttaa muotoaan ja emittoi samalla fotonin. Bioluminesenssi on kemiluminesenssin erikoistapaus. ElektroluminesenssiElektroluminesenssi syntyy kaasussa, joka on voimakkaassa sähkökentässä. Se esiintyy muun muassa salamissa, loistelampuissa ja Pyhän Elmon tulessa. TriboluminesenssiTriboluminesenssi aiheutuu eräiden materiaalin fysikaalisesta ärsyttämisestä, kuten venyttämisestä, hankaamisesta ja rikkomisesta. SonoluminesenssiSonoluminesenssi syntyy joissakin orgaanisissa nesteissä, kun ne altistetaan ultraäänelle. |
Eliöt käyttävät bioluminesenssia hyvin erilaisiin tarkoituksiin. Joillekin se on keino piiloutumiseen tai vihollisen hämäämiseen, toisille taas keino vastakkaisen sukupuolen houkutteluun. Joillekin eliöryhmille bioluminesenssin merkitystä voi vain arvailla.
Yleensä eläimet piiloutuvat väriä vaihtamalla, mutta myös valon avulla voi piiloutua: Saalistajat havaitsevat lähellä merenpintaa uivat kalat tummina silhuetteina ylhäältäpäin tulevaa valoa vasten. Helmiäiskaloilla (Argyropelecus) on vatsapuolellaan kaksi riviä valoelimiä, joiden tuottama valo on lähes yhtä kirkasta kuin veden läpi suodattunut auringonvalo. Nämä valot tekevät silhuetin selvästi vaikeammin havaittavaksi. [1] Myös eräät mustekalat ovat kehittäneet vastaavan menetelmän, mutta ne loistavat koko ruumiillaan.
|
|
|
Kuva 1: Syvänmerenhelmiäiskala (Argyropelacus aculeatus) sivulta (vasemmalla) ja alhaaltapäin (oikealla). Valoelinten tuottama valo auttaa kalaa sulautumaan veden pinnan läpi kuultavaan auringon valoon. |
Loistavien eliöiden määrä lisääntyy selvästi syvemmälle mentäessä. Monet syvänmerenkalat joutuvat tuottamaan valoa nähdäkseen, mutta sen tarkoituksena voi olla myös esimerkiksi saaliin houkutteleminen.
Syvänmerenkrotti on yksi erikoisimmista syvänmerenkaloista. Se ui koko elämänsä ajan suu auki odottaen, että sen valoelimen houkuttelema saalis uisi sisään. Krotilla on pitkät hampaat, jotka ovat tavallisesti kääntyneet sisäänpäin helpottaen näin saaliin sisäänpääsyä. Kun saalis ui sisään, hampaat loksahtavat pystyasentoon sulkien saaliin kalterien taakse. Aikuiset Krotit syövät yleensä vain kerran elämässään, ja syötyään ne ovat valmiita tuottamaan jälkeläisiä.
Suurin osa meressä esiintyvästä bioluminesenssista on väriltään sinivihreää, sillä merivesi suodattaa muut aallonpituudet nopeasti pois. Samasta syystä monet eliöt aistivat vain sinistä valoa. Malacosteid-heimon kalat ovat oppineet hyödyntämään tätä ominaisuutta: Ne tuottavat silmien alapuolella sijaitsevissa valoelimissään punaista valoa. Vaikka punainen valo ei etenekään merivedessä pitkälle, se auttaa kuitenkin kalaa näkemään saaliseläimet niiden huomaamatta. Lisäksi näillä kaloilla on erillinen valoelin, jossa ne tuottavat tavallista sinivihreää valoa.
Malacosteid-heimon kalojenkin valo syntyy sinisenä, mutta niiden valoelimissä on fluoresoivaa proteiinia, joka absorboi sinisen valon ja emittoi punaista. Tämä ei kuitenkaan vielä riitä, vaan kalojen on pystyttävä myös näkemään lähettämänsä valo. Malacosteus-suvun kaloilla on silmässään toisenlaista proteiinia, joka muuttaa valon takaisin siniseksi, jolloin ne voivat havaita sen tavallista sinistä pigmenttiä sisältävillä aistinsoluillaan. Suku Aristostomias näyttää ratkaisseen ongelman tavallisemmalla tavalla, ja kehittäneen itselleen punaista pigmenttiä.
|
|
Kiiltomadot (Lampyridae) ovat pieniä noin 10 – 20mm pituisia kovakuoriaisia, joihin kuuluu parituhatta lajia. Suomessa elää melko yleisenä Lampyris noctiluca eli ns. eurooppalainen kiiltomato. Eniten kiiltomatoja esiintyy keskikesällä aukeiden reunoilla, mutta niiden löytäminen on helpompaa elokuussa, jolloin iltayöstä kiiltomatojen parhaaseen lentoaikaan on jo riittävän pimeää. Kaikkein voimakkaimmin loistavat siivettömät noin 20 mm pitkät naaraat, jotka kiipeävät heinänkorsiin houkuttelemaan lentäviä runsaan kymmenen millimetrin mittaisa koiraita luokseen. Kiiltomatojen valo syntyy vatsapuolella sijaitsevassa kolmen läpikuultavan solukerroksen muodostamassa ns. valoelimessä, jonka naaras kääntää ylöspäin koirasta houkutellessaan. Myös koiraat, naarasta muistuttavat toukat ja jopa munat loistavat, vaikkakin naarasta heikommin. Sekä toukat että aikuiset syövät maakotiloita. Toinen Suomessa elävä kiiltomatolaji, Pikkukiiltomato (Phosphaneus hemipterus), on hyvin harvinainen, ja sen elintavat tunnetaan huonosti.
|
|
Kohtalokas myrkkyPariuduttuaan erään tulikärpäsen, Photuris pennsylvanican naaras muuttaa välähdyskuviotaan, ja alkaa vastata Photinus pyralis -koiraiden signaaleihin. Kun hämääntynyt koiras laskeutuu naaraan viereen paritellakseen, naaras tappaa koiraan ja syö sen. Tämä johtuu mitä ilmeisimmin P. pyraliksen sisältämästä Bufagiiniksi kutsutusta vaarallisesta myrkystä, joka suojaa tulikärpästä mm. lintuja ja hämähäkkejä vastaan. Syömällä yhden P. pyraliksen P. pennsylvanica saa sekä itselleen että munilleen myrkyn antaman suojan. P. pyraliksen oma myrkky voi siis koitua sen kohtaloksi. Sama myrkky on tappanut yhdysvalloissa myös lukuisia lemmikkiliskoja. Tulikärpäsiä on helppo pyydystää, ja siksi niitä onkin syötetty monille kaikkiruokaisille hyönteissyöjäliskoille, joiden alkuperäisellä elinalueella ei elä tulikärpäsiä, eivätkä ne siksi osaa niitä varoa. Lusibufogiini muistuttaa rakenteeltaan sormustinkukan myrkkyjä, digitalisglykosideja, joita oikein annosteltuna voidaan käyttää sydänlääkkeenä. Jo yksi tulikärpänen sisältää tappavan annoksen. [13, 19]
Kuva 3: Tämä yhdysvalloissa erittäin yleinen tulikärpänen (Photinus ignitus) sisältää tarpeeksi myrkkyä kookkaan lemmikkiliskon liskon tappamiseen. SynkronismiVoisi kuvitella. että tulikärpästen määrätty vastausaika johtaisi helposti synkronismiin usean naaraan vastatessa samaan väläykseen. P. pyraliksella tämä on kuitenkin hyvin harvinaista, vaikka ilmiö onkin onnistuttu tuottamaan keinotekoisesti. Eräiden aasialaisten tulikärpäslajien koiraat sitä vastoin suorastaan pyrkivät synkronismiin. Ne kerääntyvät illalla puihin, ja alkavat väläytellä valoaan tasaisin välein. Alkuillasta välähdykset sattuvat täysin epätahdissa, mutta illan edetessä ne muodostavat yhä laajempia synkronoituja ryhmiä, kunnes koko puu välkkyy tahdistetusti. Viereisissä puissa väläykset ovat myös keskenään synkronissa, mutta eri tahdissa toisen puun kanssa. [] Näillä tulikärpäsillä välkkyminen alkaa uudestaan joka ilta, mutta esimerkiksi Colophotia- suvun koirailla se saattaa pysyä muuttumattomana viikkokausia häiriötekijöistä riippumatta. [1] |
Pohjois-Amerikassa ja Etelä-Euroopassa elää tulikärpäsiksi kutsuttuja samaan heimoon kuuluvia hyönteisiä, jotka ovat kehittäneet paljon monimutkaisempia viestintämenetelmiä. Esim. pohjoisamerikkalaisen Photinus pyraliksen koiras lentää ja väläyttää valoaan säännöllisin väliajoin (n. 5,8s [1]). Kun maassa odottava naaras näkee valon, se väläyttää takaisin noin kahden sekunnin kuluttua (lämpötilasta riippuen), jolloin koiras lähtee lentämään naarasta kohti väläyttäen hetken kuluttua uudestaan, johon naaras taas vastaa. Tämä ehtii yleensä tapahtua useita kertoja ennen kuin koiras pääsee naaraan luo.
Paikalla on kuitenkin myös muita koiraita. Kuinka koiras välttää erehdyksen? On havaittu, että koiras tunnistaa naaraan vastaukseen kuluvasta ajasta, ja erehdys voi tapahtua vain, jos toinen koiras sattuu väläyttämään valoaan täsmälleen kahden sekunnin kuluttua, mutta tällöinkin suunta korjaantuu heti seuraavan välähdyksen yhteydessä. Välähdysten välinen aika kuten myös välähdyksen muoto (kuva) ja aallonpituus ovat lajikohtaisia.
Luminesenssi syntyy, kun virittyneessä tilassa oleva atomi tai molekyyli palaa takaisin normaalitilaan. Elektroni, joka molekyylin virittyessä on siirtynyt ulommalle kuorelle, palaa takaisin alemmalle kuorelle*), jolloin ylimääräinen energia vapautuu valona. Viritystilan saavuttamiseen tarvitaan kuitenkin paljon energiaa. Tarpeeksi suuren energiamäärän vapautuminen yhden reaktioaskelen aikana on hyvin harvinaista*). Bioluminesenssin mahdolliseksi syntymekanismiksi on tarjottu kahta vaihtoehtoa [12]:
1. Elektroninsiirtoreaktiossa negatiivisesti varautunut ioni luovuttaa positiiviselle ionille elektronin. Tällöin saattaa vapautua tarpeeksi energiaa jälkimmäisen virittymiseen. Välittömästi tämän jälkeen viritystila purkautuu ja energia vapautuu valona.
|
Hapen välttämättömyyden bioluminesenssille voi todeta esimerkiksi laittamalla luminoivia bakteereita tai mesisienen rihmastoa purkkiin ja pumppaamalla sen tyhjäksi. Valo himmenee ilman ohetessa ja sammuu lopulta kokonaan. Kun astiaan taas päästetään ilmaa, syntyy salaman välähdystä muistuttava äkillinen valonväläys, kun kaikki hapenpuutteen aikana syntynyt lusiferiini hapettuu kerralla. Tämän jälkeen valon voimakkuus palaa normaalille tasolle. |
2. Tarvittava energia voi myös vapautua neliatomisen peroksidirenkaan hajotessa. Peroksidirenkaassa on kaksi toisiinsa yksinkertaisilla sidoksilla liittynyttä happiatomia sekä kaksi hiiltä. Renkaan hajotessa muodostuu yleensä hiilidioksidia ja jäljelle jää esteriryhmä. Voimakasta näkyvää valoa tuottava bioluminesenssi näyttäisi olevan yhdistelmä näistä kahdesta: molekyylin sisällä tapahtuu elektronin siirto, joka laukaisee peroksidirenkaan hajoamisen. [12]
Näkyvän bioluminesenssin voi yleistää reaktioksi, jossa lusiferiiniksi kutsuttu aine hapettuu lusiferaasi-entsyymin vaikutuksesta oksilusiferiiniksi, joka virittyy. Suurin osa kemiallisesta energiasta vapautuu valona viritystilan purkautuessa, ja vain pieni osa muuttuu lämmöksi reaktion aikaisemmissa vaiheissa.
Erilaisten lusiferiinien suuresta määrästä voi päätellä, että bioluminesenssi on syntynyt evoluutiohistorian aikana itsenäisesti noin 30 kertaa[12]. Lähes kaikille bioluminesenssireaktioille näyttää kuitenkin lusiferiinin ja lusiferaasin lisäksi olevan yhteistä happimolekyylien osallistuminen reaktioon sekä läheinen yhteys soluhengitykseen.
Lusiferaasin emäsjärjestys on erilainen kaikilla eliöillä, mutta lusiferiinejä lienee yhteensä vain joitakin kymmeniä, joista ainakin seitsemän rakenne tunnetaan tarkasti[12]. Osittain tämä johtunee siitä, etteivät kaikki eliöt itse tuota bioluminesenssiin tarvittavia kemikaaleja. Monien lajien valoelimissä elää symbioottisia bakteereita, toiset taas syövät valoa tuottavia eliöitä, joiden lusiferiinin tai lusiferaasin ne käyttävät omaan bioluminesenssiinsa. Evoluutio ei myöskään mahdollista lusiferiinin rakenteen muuttumista kovin helposti, sillä monilla lusiferiineilla jopa konformaatioisomeereiltä puuttuu kyky tuottaa valoa.
|
MEDKNILYGP EPFHPLADGT AGEQMFYALS RYADISGCIA LTNAHTKENV LYEEFLKLSC
RLAESFKKYG LKQNDTIAVC SENGLQFFLP LIASLYLGII AAPVSDKYIE RELIHSLGIV
KPRIIFCSKN TFQKVNLVKS KLKYVETIII LDLNEDLGGY QCLNNFISQN SDINLDVKKF
KPNSFNRDDQ VALVMFSSGT TGVSKGVMLT HKNIVARFSH CKDPTFGNAI NPTTAILTVI
PFHHGFGMTT TLGYFTCGFR VALMHTFEEK LFLQSLQDYK VESTLLVPTL MAFFPKSALV
EKYDLSHSKE IASGGAPLSK EIGEMVKKRF KLNFVRQGYG LTETTSAVLI TPDTDVRPGS
TGKIVPFHAV KVVDPTTGKI LGPNETGELY FKGDMIMSKY YNNEEATKAI INKDGWLRSG
DIAYYDNDGH FYIVDRLKSL IKYKGYQVAP AEIEGILLQH PYIVDAGVTG IPDEAAGELP
AAGVVVQTGK YLNEQIVQNF VSSQVSTAKW LRGGVKFLDE IPKGSTGKID RKVLRQMFEK
HKSKL
Kuva 7: Erään pohjoisamerikkalaisen tulikärpäsen (Photuris pennsylvanica) 545 aminohappoa sisältävän lusiferaasin aminohappojärjestys. [26] |
|
|
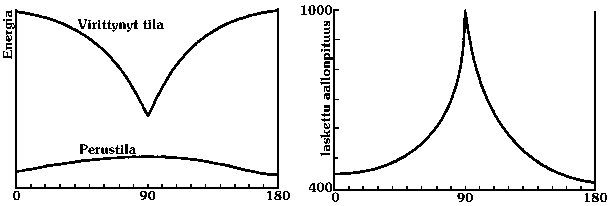
VärivaihteluaBioluminesenssin väri vaihtelee merkittävästi eri kiiltomato- ja tulikärpäslajien välillä, vaikka ne käyttävät samaa lusiferiinia. Valon aallonpituus riippuukin pääasiassa jokaiselle lajille ominaisesta lusiferaasin rakenteesta. Lisäksi ulkoisten tekijöiden kuten lämpötilan, matalan pH:n tai metalli-ionien on todettu voivan nostaa aallonpituutta normaalista vihreästä (552 nm – 528 nm) jopa punaiseen (noin 610 nm).[4] Kiiltomadon oksilusiferiinin aktiivinen osa on tavallisesti samassa tasossa muun rungon kanssa. Se pääsee kuitenkin helposti kiertymään sitä muuhun runkoon yhdistävän hiili-hiili-sidoksen ympäri siten, että molekyylin energia kasvaa vain vähän, sillä sidoksen lähellä ei ole kiertymistä estäviä vetyatomeita. Kun lusiferiini on sitoutuneena lusiferaasi-entsyymiin, entsyymin vetyatomit muodostavat vetysidoksia lusiferiinin typpi- ja rikkiatomeiden kanssa ja pakottavat näin sen aktiivisen kohdan tiettyyn kulmaan muuhun runkoon nähden. Lusiferiinin virittyessä tilanne kuitenkin muuttuu. Tällöin eri kiertokulmia vastaavat molekyylienergiat eivät olekaan enää lähellä toisiaan, vaan 90 asteen kiertokulma osoittautuu selvästi vähäenergisimmäksi. (Tavallisesti molekyyli asettuu heti pienienergisimpään tilaansa, mutta tässä tapauksessa fotoni emittoituu ennen kuin molekyylin konformaatio ehtii muuttua.) Kuten koulukursseista on tuttua, viritystilan purkautuessa vapautuvan valon aallonpituus vastaa virittyneen ja normaalitilaisen molekyylien välistä energiaeroa. Kun molekyylienergia riippuu merkittävästi kiertokulmasta, vaikuttaa kulma vastaavasti myös fotonin aallonpituuteen. [2] Tietokoneella laskettu molekyylienergioiden ja aallonpituuksien riippuvuus kiertokulmasta käy ilmi seuraavista kaavioista:
|
Kiiltomatojen bioluminesenssireaktiossa tarvittava energia saadaan adenosiinitrifosfaatista (ATP).
LH2+ATP+O2
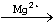 P+AMP+PPi+CO2+hv
P+AMP+PPi+CO2+hv
ATP hajoaa pyrofosfaatiksi (PPi) ja AMP:ksi, joista jälkimmäinen reagoi lusiferiinin (LH2) kanssa muodostaen lysiferyyliadenylaattimolekyylin. Molekyylin aktiiviseen kohtaan muodostuu peroksidirengas, joka hajoaa hiilidioksidiksi ja karbonyyliryhmäksi vapauttaen energiaa. Näin syntynyt oksilusiferiini (P) virittyy ja emittoi fotonin viritystilan purkautuessa. (ks. kuva 4) Lusiferaasi katalysoi sekä lusiferyyliadenylaatin muodostumista että peroksidirenkaan hajoamista. Lusiferaasin lisäksi reaktio edellyttää magnesiumionien läsnäoloa. Emittoidessaan fotonin oksilusiferiini on vielä kiinni lusiferaasissa. [12]
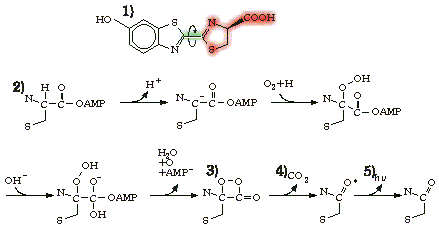
|
|
Kuva 5: Kiiltomatojen lusiferiini (1) (Sidos, jonka ympäri kiertyminen tapahtuu (kts. laatikko s. ) on merkitty kiertävällä nuolella ja aktiivinen osa punaisella taustalla) sekä sen oletettu lusiferaasin katalysoima hapettumisreaktio (vain aktiivinen osa näkyvissä): AMP kiinnittyy lusiferiiniin(2). Lusiferiinin aktiiviseen kohtaan muodostuu peroksidirengas (3), joka hajoaa karbonyyliryhmäksi ja hiilidioksidiksi (4). Reaktiossa vapautuu niin paljon energiaa, että karbonyyliryhmän happiatomi jää virittyneeseen tilaan. Viritystilan purkautuessa syntyy valoa (5). (Reaktioyhtälö on muodostettu lähteiden [7, 12] perusteella.) |
Kiiltomatojen bioluminesenssireaktio on yksi tehokkaimpia. Sen hyötysyhde on noin 0,92 eli 92% energiasta vapautuu valona. Ideaalisissa olosuhteissa fotoneja saattaa syntyä jopa yksi jokaista hapettunutta lusiferiinimolekyyliä kohden, mutta yleensä tuotanto ei ole näin hyvä.
Luminoivia bakteereita elää kaikkialla meressä. Ne tuottavat valoa kuitenkin vain tiheissä kasvustoissa, ja niiden loisteen voi havaita esimerkiksi mädäntyneessä kalassa, jossa bakteerit ovat päässeet kasvamaan tarpeeksi runsaslukuisiksi. Bakteereista parhaiten tunnettuja ovat meressä elävät Vibrio harveyi, V. fischeri, Photobacterium phosphoreum ja P. leiognathi sekä maalla elävä Xenorhabdus luminescens [17].
Bakteerien bioluminesenssi on erityisen läheisessä yhteydessä soluaineenvaihduntaan, sillä niiden lusiferiinina toimii pelkistynyt flaviinimononuklidi (FMNH2), joka tavallisesti toimii vedyn siirtäjänä. [10, 18] Bakteereissa tapahtuva lusiferaasin katalysoima luminesenssireaktio voidaan yleistää seuraavasti:
FMNH2 + RCHO ® FMN + RCOOH + H2O + hv (l = 490nm)
missä RCOOH on pitkäketjuinen alifaattinen aldehydi, jonka hapettuminen kuuluu välttämättömänä osana luminesenssireaktioon. [10, 17, 18]
Monet suuremmat eliöt elävät symbioottisessa suhteessa valaisevien bakteerien kanssa. Näillä on ihanteelliset isännän kasvuolosuhteet valoelimessä. Erään kalmarin (Euprymna scolopes) valoelimissä elää Vibrio fischeri -bakteereita. Näitä bakteereita on kaikkialla merivedessä noin 100 kappaletta millilitrassa, mutta tällöin ne eivät kuitenkaan tuota valoa. Valon tuotanto alkaa vasta, kun bakteeritiheys nousee 1010 – 1011 soluun/ml kuten esimerkiksi valoelimissä.
Koska V. fischerin valontuotanto liittyy symbioottiseen suhteeseen, se on turhaa bakteerien eläessä vapaana merivedessä tai vasta kehittymässä olevissa valoelimissä, joiden bakteerikanta ei muutenkaan olisi tarpeeksi vahva tuottamaan merkittävää valomäärää. Niinpä bakteerit ovatkin kehittäneet erityisen keinon muiden bakteerien läheisyyden aistimiseksi.
Bakteerit tuottavat autoindusoijaksi kutsuttua molekyyliä, joka rakenteensa vuoksi pystyy vapaasti kulkemaan solukalvon läpi. Kun bakteereita on harvassa, autoindusoija diffundoituu ympäröivään veteen, mutta tiheissä kasvustoissa solujen välinen tila alkaa täyttyä, ja autoindusoijaa siirtyy yhä enemmän takaisin solujen sisälle.
Kaikkien V. fischerin luminesenssigeenien toimintaa säätelee luxR:ksi kutsuttu proteiini, jota on jatkuvasti solun solulimassa noin 500 molekyyliä. luxR:ssä on kaksi aktiivista kohtaa, joista toinen reagoi autoindusoijan ja toinen DNA:n lux-operonin kanssa. Kun autoindusoija kiinnittyy luxR:ään, se aktivoituu ja käynnistää luminesenssiin tarvittavien proteiinien tuotannon. Samalla syntyy lisää autoindusoijaa, joten sen määrä kasvaa entisestään. [17]
Symbioottisten bakteerien tuottaman valon säätely on yleensä mekaanista: Valon pääsy ulos valoelimistä estetään esimerkiksi suljinläpällä. Eräillä eliöillä symbioottiset bakteerit elävät valoelimen solujen sisällä eli ovat evolvoitumassa soluelimiksi*). Tällöin eliöt voivat säädellä valonvälähdyksiä valoelinsolujen lähettämien ärsykkeiden avulla. Tarkkaa säätelymekanismia ei kuitenkaan tunneta. [12]
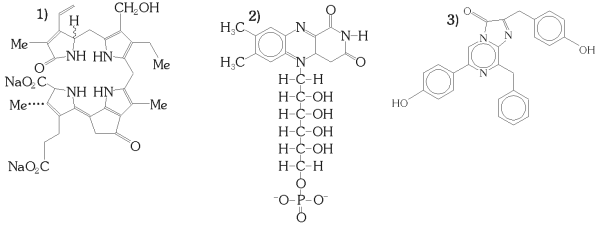
Kuva 9: Erilaisia Lusiferiineja: 1) Panssarisiimaeliöiden lusiferiini, jonka uskotaan olevan klorofyllin johdannainen. Sen hapettumista estävä lusiferiininsitojaproteiini reagoi pH:n muutoksiin. 2) Bakteerien lusiferiini on pelkistynyt flaviinimononukleotidi (FMNH2), joka tavallisesti toimii soluaineenvaihdunnassa vedyn siirtäjänä. Joissakin bakteereissa se osallistuu bioluminesenssireaktioon hapettuessaan yhdessä erään pitkäketjuisen alfaattisen aldehydin kanssa. 3) Coelenterasiini, joka osallistuu mm. aequoriinin muodostamiseen. |
|
Viequesin saarella Karibianmerellä sijaitsee Mosquito Bayn nimellä tunnettu lahti. Kun lahdelle saapuu yöllä, on aivan pimeää, mutta pienikin kosketus vedenpintaan aiheuttaa voimakkaan valonvälähdyksen. Loiste aiheutuu pienistä panssarisiimaeliöistä, joita vedessä on noin 133 000 kappaletta litrassa. Lahti on aikaisemmin ollut suosittu turistikohde, mutta nykyään se on rauhoitettu, ja sinne menemiseen tarvitaan erityislupa. Ilmansaasteet ovat kuitenkin jo häirinneet lahden herkkää ekosysteemiä, ja sen kirkkaus on nykyään vain kymmenesosa alkuperäisestä. [10] |
Panssarisiimaeliöt ovat pieniä vedessä keijuvia eliöitä, jotka mekaanisesti ärsytettyinä synnyttävät valonväläyksiä.
Hankajalkaiset ovat panssarisiimaeliöiden luonnollisia vihollisia, mutta siimaeliöt ovat kehittäneet ainutlaatuisen puolustusmekanismin: Kun hankajalkaiset lähestyvät siimaeliöparvea, ne aiheuttavat veteen virtauksia, jotka laukaisevat siimaeliöiden bioluminesenssin. Valo puolestaan houkuttelee paikalle kaloja, jotka syövät hankajalkaiset pois häiritsemästä. Näin kalat ja panssarisiimaeliöt hyödyttävät toinen toisiaan.
Monien muiden eliöiden tavoin panssarisiimaeliöiden bioluminesenssi on läheisessä yhteydessä soluaineenvaihduntaan ja fotosynteesiin. Niiden tuottaman valon voimakkuus riippuu auringonvalon määrästä edellisenä päivänä. Mitä enemmän aurinkoa ne ovat saaneet, sitä kirkkaammin ne loistavat. Panssarisiimaeliöiden lusiferiinikin muistuttaa rakenteeltaan klorofylliä, jonka johdannainen sen arvellaankin olevan.
Panssarisiimaeliöt säätelevät bioluminesenssireaktiota nk. lusiferiininsitojaproteiinin avulla. Normaali-pH:ssa (8) proteiini sitoutuu lusiferiiniin estäen näin sitä reagoimasta, mutta pH:n laskiessa kuuteen proteiini menettää toimintakykynsä, jolloin luminesenssireaktio käynnistyy. Valo syntyy 0,1 sekunnin pituisina välähdyksinä, joiden aikana eliöstä lähtee noin 6×108 fotonia. Valo on niin kirkas, että jopa yhden eliön tuottaman valon voi helpoasti havaita pimeään tottuneella silmällä. [10]
Meressä elävillä eliöillä yleisin lusiferiini on nk. coelenterasiini. Se esiintyy lukuisissa eläinryhmissä, kuten kaloissa ja kalmareissa. Eri eliöt soveltavat coelenterasiinia eri tavoin. Joillakin se osallistuu tavalliseen hapettumisreaktioon, mutta toisilla se reagoi lusiferaasin ja hapen kanssa muodostaen suuren fotoproteiinikompleksin, jota kutsutaan aequoriiniksi. Valoa syntyy kalsium-ionin (Ca2+) osuessa fotoproteiiniin. Tämä on ainoita tunnettuja bioluminesenssireaktioita, jotka voivat tapahtua anaerobisesti eli ilman happimolekyylin välitöntä läsnäoloa. Vaikka happea tarvitaankin aequoriinin muodostamiseen, aequoriini voi säilyä hyvinkin pitkään reaktiokykyisenä mahdollistaen näin bioluminesenssin silloinkin kun happea ei ole saatavilla.
Bioluminesenssin evoluution ymmärtäminen ovn monessa suhteessa ongelmallista. Valon tuotanto kuluttaa runsaasti energiaa, ja siksi sen ylläpitäminen on lajin säilymisen kannalta epäedullista, ellei siitä ole jotakin muuta merkittävää hyötyä. Jotta jokin hyödyllinen ominaisuus voisi syntyä, on myös kaikkien välivaiheiden oltava lajin säilymiselle edullisia. Luonnollisesti lusiferiinin ja lusiferaasin esiasteiden tuottaminen on hengissä selviytymisen kannalta vain energian tuhlausta niin kauan kuin valoa ei synny, ja tarpeeksi monen mutaation sattuminen yhtä aikaa on äärimmäisen epätodennäköistä. Siksi onkin oletettavaa, että bioluminenenssi on aluksi syntynyt jonkin muun, ehkä sittemmin tarpeettomaksi käyneen reaktion sivutuotteena. Monille lusiferaaseille onkin löydetty vastineita solun normaaliin aineenvaihduntaan osallistuvista entsyymeistä, ja bioluminesenssireaktiot ovat muutenkin usein läheisessä yhteydessä soluhengitykseen.
On ehdotettu, että bakteerien bioluminesenssireaktio olisi alunperin kehittynyt hapen poistamiseen. Vaikka molekylaarinen happi on monille eliöille elintärkeää, se on itse asiassa erittäin vaarallista niillekin soluille, jotka ovat sopeutuneet käyttämään sitä energialähteenään. Ihminenkin voi kuolla hapen yliannostukseen.
|
|
Lahokannoissa kasvavan mesisienen rihmaston tuottaman heikon valon voi nähdä pimeänä syyskesän öinä, kun menee metsään ilman taskulamppua ja antaa silmiensä tottua pimeään. Monien japanilaisten sienten itiöemätkin loistavat varsin kirkkaasti, ja ne on helppo nähdä metsässä pitkänkin matkan päästä.
Bioluminesenssin merkitys sienille tunnetaan hyvin huonosti. On ehdotettu, että valo houkuttelisi hyönteisiä itiöiden levittäjiksi, mutta tämä teoria ei sovi ainakaan mesisieneen. Todennäköisempää onkin, että bioluminesenssi on jonkinlainen aineenvaihdunnan sivutuote.
Bioluminesenssi on tunnettu jo tuhansia vuosia, ja sille on kehitetty lukuisia niin käytännön elämään kuin kemianteollisuuteen ja lääketieteeseen liittyviä sovelluksia. Erilaisia bioluminesenssireaktioita voidaan käyttää jonkin niihin olennaisesti kuuluvan ainesosan indikaattoreina. Näistä tärkein on ATP-testi, jossa käytetään kiiltomatojen lusiferiinia ja lusiferaasia. ATP-testillä voidaan havaita esimerkiksi virtsassa olevia bakteereita sekä testata bakteerien vastustuskykyä antibiooteille: bakteereita viljellään petrimaljassa, ja käsitellään viljelmä testattavalla antibiootilla. ATP-testi kertoo, onko bakteereita vielä elossa[4].
Tieteellisessä tutkimustyössä tärkeimmät sovellukset liittynevät geenitekniikkaan. Koska bioluminesenssi on helppo havaita, sitä voidaan käyttää indikaattorina geenin siirtymisestä. Kun esim. V. fischeristä eristetty geeni siirretään johonkin toiseen eliöön jonkin tietyn geenin yhteyteen, tämän geenin esiintymistä geenimanipuloiduissa eliöissä voidaan seurata. Bioluminesenssigeeniä ilmentävän eliön perimä sisältää nimittäin hyvin suurella todennäköisyydellä myös seurannan kohteena olevaa geeniä.
Panssarisiimaeliöiden bioluminesenssigeenejä voidaan käyttää myös biologisen kellon tutkimiseen, sillä niiden toiminta riippuu vuorokaudenajasta. Kun geenejä siirrettiin tupakantaimiin, nekin alkoivat loistaa jaksottaisesti. Vaihtelurytmi saattoi kuitenkin normaalista vuorokausikierrosta poiketen olla esimerkiksi 12 tai 36 tuntia.
Käytännön elämässä bioluminesenssia on käytetty esimerkiksi valaisimena tai syöttinä: Japanissa sotilaat kuljettivat aikoinaan mukanaan kuivattuja äyriäisiä. Kun niitä hieroi sormiensa välissä, ne tuottivat tarpeeksi valoa kartan lukemiseen. Euroopassa puolestaan on saatettu laittaa lyhtyyn kiiltomatoja tai lahokannonpalasia kynttilänkorvikkeeksi.
Artikkelin asiasisällön on tarkistanut
Elimistön säätelyä valon nopeudellaViime vuosisadan puolivälistä lähtien suoritetuissa tutkimuksissa on havaittu, että lähes kaikki elävät organismit bakteereista ihmiseen säteilevät heikkoa joko näkyvää tai ultraviolettia valoa. Valo on niin heikkoa, että sen voi havaita ainoastaan yksittäiset fotonit laskevilla mittalaitteilla, sillä fotoneita lähtee vain 10 – 1000 kappaletta sekunnissa neliösenttimetriltä*).[3] Valoa voidaan myös vahvistaa luminoliksi kutsutun yhdisteen avulla[5]. Vaikka ilmiö tulikin yleisesti hyväksytyksi vasta 1950-luvulla, sen oli havainnut venäläinen Alexander Gurwitsch jo 1922[3]. Tällä nk. ultraheikolla bioluminesenssilla on tärkeä merkitys monissa elimistön toiminnoissa – erityisesti elimistön säätelyjärjestelmässä. Se kuljettaa monia viestejä, joiden aiemmin oli luultu siirtyvän ainoastaan hormonien välityksellä. On mm. havaittu, että kun kahdesta kvartsilevyllä toisistaan erotetulle rotalle syötettiin hormoneja, hormonituotanto käynnistyi myös toisessa rotassa. Kvartsilevyn korvaaminen lasilevyllä poisti tämän ilmiön[3]. Ultraheikko bioluminesenssi toimii myös osana elimistön puolustusjärjestelmää. Valkosolut säteilevät valoa aina bakteerin syömisen yhteydessä. Näin ne saattavat käynnistää vasta-ainetuotannon myös muissa valkosoluissa[3]. Tämä ilmiö on johtanut lukuisiin lääketieteellisiin sovelluksiin, kuten ihosyövän[3], verisyövän sekä tuberkuloosin[5] toteamiseen. Lisäksi sen avulla voidaan tutkia mm. alkion kehitystä ja siementen elinkelpoisuutta[3]. Ultraheikon bioluminesenssin synnyttävät mekanismit tunnetaan erittäin huonosti. Fotonien lähteenä on todennäköisesti molekylaarinen happi. Hapella on kaksi viritystilaa, jotka tulevat kysymykseen ultraheikon bioluminesenssin lähdettä etsittäessä. Viritystilaa, jossa yksi elektroni on siirtynyt ulommalle kuorelle merkitään 1DO2:lla. Paluu tästä viritystilasta normaalitilaan (3SO2) vapauttaa fotonin, jonka aallonpituus on 1268 nm. Hapella on toinenkin, vähempienergiainen viritystila, jossa elektronit pysyvät samalla pääkuorella, mutta sijaitsevat vastoin Hundin sääntöä samalla alakuorella. Tätä viritystilaa merkitään 1SO2, ja sen purkautuessa vapautuvan fotonin aallonpituus on 762 nm. [5] Todellisissa tilanteissa aallonpituuteen vaikuttaa paljon muitakin tekijöitä. Yleensä ultraheikon bioluminesenssin aallonpituus vaihtelee välillä 300 – 800 nm. |
Kuvia löytyy osoitteista
http://lifesci.ucsb.edu/~biolum/organism/photo.html
http://www.biobay.com/.
[1] Harvey, E. Newton; Bioluminescence. Academic Press, New York 1952
[2] Hastings, J. W.; Kricka, L. J.; Stanley, P. E. (toim.); Bioluminescence and Chemiluminescence – Molecular Reporting with Photons. John Wiley & Sons, New York 1996.
[3] Chang, Jiin-Ju; Fisch, Joachim; Popp, Fritz-Albert; Biophotons. Kluwer Academic Publishers, Dordrecht 1998.
[4] DeLuca, Marlene A.; McElroy, William D.; Methods in Enzymology Volume 133 Bioluminescence and Chemiluminescence Part B. Academic Press, Orlando 1986.
[5] Van Dyke, Knox; Castranova, Vincent; Cellular Chemiluminescence, Vol I, II ja III. CRC Press, 1987.
[6] Crowson, R. A.; The Biology of the Coleoptera. Academic Press, Lontoo 1981.
[7] Rockstein, Morris; The Biochemistry of Insects.Academic Press, Lontoo 1978
[8] Tirri, Rauno; Lehtonen, Juhani; Lemmetyinen, Risto; Pihakaski, Seppo; Portin, Petter; Biologian sanakirja. Otava, Keuruu 1993.
[9] Branham Marc (1998); The Firefly Files. http://iris.biosci.ohio-state.edu/projects/FFiles/biolum.html
[10] Haddock, S.H.D.; McDougall, C.M.; Case, J.F. (2000); “The Bioluminescence Web Page”, http://lifesci.ucsb.edu/~biolum/
[11] LLosa, Frank (2000); World’s Brightest Glowing Bay! http://www.biobay.com/
[12] Hastings, J. W.; Biological diversity, chemical mechanisms, and the evolutionary origins of bioluminescent systems / Journal of Molecular Evolution vol. 19: p.309-321. Springer–Verlag 1983.
[13] Vaarallista Lemmikinruokaa. Tieteen Kuvalehti 16/1999 p. 15, Bonnier Julkaisut Oy.
[14] Microsoft Encarta 98 Encyclopedia.
[15] Light in the Ocean: Bioluminescence. Plymouth Marine Laboratory, http://www1.npm.ac.uk/set98/set97/biolumin.htm
[16] Bioluminescence. Scripps Institution os Oceanography, http://siobiolum.ucsd.edu/Biolum_intro.html
[17] Bluth, Brian J.; Frew, Sarah E.; McNally, Brian; (³1997), Cell-Cell Communication and the lux operon in Vibrio fischeri. Department of Biological Sciences, Carnegie Mellon University, http://www.bio.cmu.edu/courses/03441/TermPapers/97TermPapers/lux/bioluminescence.html
[18] Frye, Mike (1999); Bioluminescence – A ”Glowing” Report. http://edie.cprost.sfu.ca/~rhlogan/frye.html
[19] Mimicry and murder in the firefly world, Environmental News Network, 2. 9. 1997
[20] Feynman, Richard P.; Leighton, Robert B.; Sands, Matthew; The Feynman Lectures on Physics, Part III – Quantum Mechanics. California Institute of Technology, 1965.
[21] Lavonen, Jari; Kurki-Suonio, Kaarle; Hakulinen, Harri; Galilei 8 – Moderni fysiikka. Weilin + Göös, Helsinki 1996.
[22] Cronin, T. W.; Järvilehto, M.; Weckström, M.; Lall, A. B.; Tuning of photoreceptor spectral sensitivity in fireflies (Coleoptera: Lampyridae) / Journal of Comp Physiol A (2000) 186: 1-12. Springer–Verlag.
[23] Light in the ocean: bioluminescence. Plymouth Marine Laboratory. http://www1.npm.ac.uk/set98/set97/biolumin.htm
[25] McCapra, Frank; Mechanisms of Bioluminescence and Chemiluminescence – Unfinished Business. Kirjassa [2]
[26] Leach, F. R.; Ye, L., Schaeffer, H. J.; Buck, L. M.; Cloning and Sequencing of a Firefly Luciferase from Photuris pennsylvanica. Kirjassa [2]
[27] Lloyd, James E.; Occurence of Aggressive Mimicry in Fireflies. Florida Entomologist 67(3) p. 368–378 (1984)
 Kuva 6: Erään pohjoisamerikkalaisen tulikärpäslajin (Photinus ignatus)
koiras väläyttelee valoaan.
Kuva 6: Erään pohjoisamerikkalaisen tulikärpäslajin (Photinus ignatus)
koiras väläyttelee valoaan.
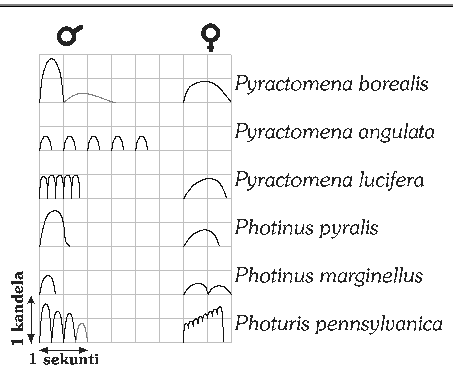 Kuva 8: Tulikärpästen välähdyskuvioita. Suomalaiset kiiltomadot
puuttuvat listasta, sillä ne tuottavat jatkuvaa valoa. Koiraan ja naaraan
välähdysten väliset aikasuhteet eivät näy kuvassa.
Kuva 8: Tulikärpästen välähdyskuvioita. Suomalaiset kiiltomadot
puuttuvat listasta, sillä ne tuottavat jatkuvaa valoa. Koiraan ja naaraan
välähdysten väliset aikasuhteet eivät näy kuvassa.